

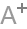
人脑结构网络的构建,可以分别基于结构磁共振图像(灰质的形态学指标, 如皮层厚度、皮层曲面积等; 蓝色箭头所示流程)和扩散磁共振图像(白质纤维束, 绿色箭头所示流程)。
人脑功能网络,可以分别基于功能磁共振图像(大脑功能活动的时间序列, 红色箭头所示流程)和脑电/脑磁信号(黄色箭头所示流程).
网络节点定义: 结构、扩散和功能磁共振数据需要利用先验图谱划分脑区或图像体素定义网络节点, 而脑电/脑磁数据则直接以记录电极/通道为网络节点.
网络连接(边)定义: 基于结构磁共振数据的网络连接定义为网络节点形态学指标之间的统计关系, 扩散磁共振数据通过确定性或概率性纤维跟踪技术确定网络节点之间的解剖连接, 基于功能磁共振及脑电/脑磁的网络连接一般可以通过皮尔森相关、偏相关、同步似然性等计算方法度量网络节点的神经活动信号之间的统计关系.
构建人脑结构和功能网络: 可以对第3 步得到的相关矩阵进行二值化, 获得不同阈值下的二值矩阵, 即大脑结构和功能网络。
下图是目前研究常用方法的流程图。
(山有小口,仿佛若有光。便舍船,从口入…)
迄今为止, 结构磁共振成像已经被大量地用于研究人脑正常发育、老化和疾病过程中局部脑区形态学的变化. 值得注意的是, 目前几项研究发现, 人脑的形态学数据中包含着大量的脑连接信息。
2005年, 英国的Mechelli 等人用结构磁共振成像发现人脑的某些脑区(如大脑的两个半球区域)之间的灰质密度具有协调的变化, 研究者推测这种协调性可能与连接它们的白质纤维束(胼胝体)有关。
2006 年,Lerch 等人发现, 大脑皮层的Broca 和Wernicke 区(2 个与语言有关的脑区)在灰质皮层厚度上具有非常高的协调性。 为此, 他们构建了一幅皮层厚度的关联图, 发现该图与人脑的弓形束( 连接Broca 和Wernicke 区的白质纤维)图谱具有惊人的相似性. 尽管目前我们还不清楚脑区间形态学相关性确切的生理意义, 但是一些研究已经表明这种形态特征的协调变化可能与先天遗传及后天的可塑性有关。
2007 年, He 等人采用124 人的结构像数据, 通过考察大脑皮层54 个脑区皮层厚度之间的相关性成功地构建了第一个人脑结构网络, 并发现该网络具有“小世界”特性, 其节点度分布服从指数截尾的幂律分布. 该研究首次提出了用形态学指标构建大脑结构连接组的思想, 并证实了由大脑皮层厚度相关性构建的人脑结构网络具有“小世界”属性, 为描述活体人脑的结构连接网络提供了一种新的途径。
2008年, Chen 等人进一步发现, 人脑皮层厚度网络具有对应于人脑功能模块(比如语言、记忆及视觉等)的组织模式, 表明了皮层厚度的关联变化主要位于不同的功能模块内. 此外, 通过比较网络中节点和边的介数(betweenness),发现人脑皮层厚度网络的核心节点主要分布在大脑顶叶, 颞叶和额叶的联合皮层区域, 而网络中的大多数重要路径则连接着不同模块中的核心节点。
2008 年, Schmitt 等人建立了600个儿童被试大脑结构的遗传相关矩阵, 通过网络分析发现, 由遗传因素调控的不同脑区之间的结构连接构成了一个具有“小世界”属性的复杂网络, 该网络中的核心脑区主要分布在额上回、额中回、中央前回及中央后回等区域. Lenroot 等人用He 等人提出的方法研究了787 个健康被试的纵向结构数据,探讨了大脑发育对脑结构(皮层厚度)网络拓扑属性的影响. 该套数据按照年龄划分为3 组, 即儿童组(平均年龄6.9 岁)、少年组(平均年龄11 岁)和青少年组(平均年龄16.4 岁), 分析发现3 个年龄组的被试大脑结构网络都具有“小世界”属性, 而且随着年龄增长,网络的聚类系数和等级特性都显著增大, 表明大脑的发育与大脑结构的区域分化密切相关。
表1 列举了基于结构磁共振成像的脑结构网络领域中研究文献
上述基于结构磁共振数据的研究表明, 大脑结构网络具有“小世界”属性和模块化结构等拓扑属性.基于形态学数据构建大脑结构网络的方法为描述人脑的结构连接模式提供了一种简单而有效的新途径,这不仅有助于揭示人脑结构网络的拓扑模式, 也为探索人脑结构与功能的相互关系提供了实验依据。
需要注意的是, 这种基于形态学数据构建大脑结构网络的方法也存在一些局限性. 比如, 目前脑形态学的结构网络通常是对全脑脑区(如N<200)间连接模式的描述, 当脑网络被扩展到体素水平(如N>10000)时,上述的基于脑区水平的网络属性是否还能被发现呢?这些不同水平的脑网络之间又存在着怎样的关系?
此外, 该方法目前只能基于人群得到一个大脑结构网络, 而无法针对单个被试建立基于个体的脑结构网络, 这使得该方法的应用具有一定的局限性, 尤其是在临床诊断研究等方面。
周末清早,天气太好,多多加班,少往外跑!
明天见。
感谢/brain.bnu.edu.cn。



